
Half a mile below the surface of the ocean, off the coast of Oregon, the Alvin submersible's headlights flicker on to reveal a colorful oasis. Plush carpets of white, yellow and orange microbes cover the seafloor, punctuated by fields of clams and mussels. Red rockfish watch the vessel warily with bulbous milky eyes, while bubble plumes belch from mounds of chalky, variegated rock. The halo of illumination draws visitors forward like a lure, exposing this alien terrain bit by unexpected bit while obscuring its true extent.
Hours earlier on this expedition in 2010, one of us (Marlow) had wriggled his way into Alvin's titanium sphere, along with two other explorers. We pressed our faces to the circular windows as we descended through a kaleidoscope of blue. Our destination was Hydrate Ridge, a rocky precinct where vast quantities of methane are being squeezed out of Earth's crust. With the accelerating pace of discovery of such methane seeps, as they are known (450 were found during a single 2016 expedition in the eastern Pacific), scientists are racing to understand their environmental impact. Methane, after all, is a strong greenhouse gas: although it constitutes only 0.00018 percent of the atmosphere, it accounts for 20 percent of the atmosphere's overall warming potential. Estimates suggest that roughly 10 percent of atmospheric methane emerges from seafloor seeps every year. Unchecked, this bubble stream could wreak climate havoc, but something prevents more methane from reaching the atmosphere: the microbes living in the seeps.
These microbes, which dwell underneath the white microbial mats and clam shards, consume methane with remarkable voracity. Individually minuscule but collectively mighty, they work together in ways that help to shape landscapes, sustain ecosystems and impact the planet's climate. Their power lies in their cooperation. Scientists have known about these microorganisms for decades, yet they remain mysterious in many respects. Key among the unknowns is the extent of their influence: Do they reside in only a few regions of the ocean floor, or are they widespread? More broadly, is their propensity for cooperation exceptional among microbes, or is it commonplace? Prevailing views long held that such organisms mostly compete with one another for resources. But maybe teamwork is actually their default mode. We were there—a speck of light suspended in the inky expanse—to figure out just how pervasive this way of life really is.
On supporting science journalism
If you're enjoying this article, consider supporting our award-winning journalism by subscribing. By purchasing a subscription you are helping to ensure the future of impactful stories about the discoveries and ideas shaping our world today.
A Microbial World
In a sense, our journey to collect microscopic organisms from the deep sea was a logical step in the broader scientific quest to understand how our planet works—how elements such as carbon, nitrogen, sulfur and phosphorus move between ecosystems or how greenhouse gases enter the atmosphere. We live, after all, in a microbial world: from rocks deep below the seafloor to desert dust particles high up in the atmosphere, microbes exist almost everywhere we look. And scientists have long recognized that they perform important roles in distributing these elements and compounds in ways that help make Earth the planet it is today, conveniently habitable for animals like us.

Credit: Tami Tolpa; Sources: “A Marine Microbial Consortium Apparently Mediating Anaerobic Oxidation of Methane,” by Antje Boetius et al., in Nature, Vol. 407; October 5, 2000; “Single Cell Activity Reveals Direct Electron Transfer in Methanotrophic Consortia,” by Shawn E. McGlynn et al., in Nature, Vol. 526; October 22, 2015; “Methane-Derived Carbonates and Authigenic Pyrite from the Northwestern Black Sea,” by J. Peckmann et al., in Marine Geology, Vol. 177, Nos. 1–2; June 30, 2001
But the approach that researchers have typically taken to studying the microbial world has limited their understanding of these globally relevant processes. For decades investigators focused their attention on individual species and their molecular components. From the teeming microbial masses between grains of sand, they isolated single organisms and poked and prodded them to suss out their biochemistry and the functions of their genes. This method has produced reams of information about these species, as well as how cells and biomolecules work in general. When researchers then zoomed out from these studies to understand the biosphere as a whole, stitched together from the constituent species, however, major gaps in their knowledge remained. Only a tiny fraction of microbes seen in the wild could be isolated, suggesting that the species making up complex natural communities are intertwined in ways that cannot easily be replicated in the laboratory. And the very coexistence of many thriving species, which often play complementary roles, seemed to contradict the conventional wisdom that microbial ecosystems revolve around a winner-take-all struggle for resources.
Moreover, metabolic activity rates of individual species measured in the lab—such as how quickly they produce oxygen or consume nitrogen—rarely matched values from real-world environments because species that can be isolated in the lab are often more vigorous than those that cannot be. In other words, the whole was sometimes more, sometimes less, but always different from the sum of the parts.
But a growing body of evidence suggests that these disconnects can be reconciled by considering the vital importance of interactions among organisms. Over the past decade advances in biomolecular sequencing and microscopic imaging, among other technologies, have enabled researchers to study microbial communities more holistically than ever before. The latest findings indicate that collaboration is a critical driver of the biosphere: as individual organisms evolve to share energy, genetic information and metabolic duties, they unlock new ways of life and gain entry to previously inaccessible habitats.
Hiding in Plain Sight
Back at Hydrate Ridge, Alvin's robotic arm plunges a clear plastic tube with an open bottom into a wispy microbial mat. It slides down easily at first but then catches, the resistance propagating back to the submersible and delivering an unexpected jolt. With a final push, the tube punches through the stubborn layer and obtains a sample, which trails a fine plume of sedimentary dust as Alvin's arm carries it to the sub's quiver of tubes.
Later that afternoon, in the ship's expansive lab, Marlow and his colleagues examine the foot-deep cross section of the seafloor that we recovered. Under the white mat, beige mud transitions into black goo and chunks of rock—the crust that briefly resisted our sampling effort—and finally tapers off to a dark gray mixture. Our microbial quarry inhabits the darkest layer, which reeks of rotten eggs. Previous work in the 1980s had shown that this was the zone where methane produced in deeper horizons and sulfate from the overlying seawater were both being removed from the sediment. Yet efforts to identify individual microbial species in this layer that could simultaneously consume methane and sulfate came up empty again and again. Taking a different tack, other researchers used methane and sulfate as bait to lure the thief out of hiding, tracking the molecules as they disappeared from experimental treatments. Some impressive sleuthing in the early 2000s showed that the culprit was not an “it” but rather a “they”: cell clumps made of two types of microbes lit up with telltale signs of metabolic activity. One partner ate methane; the other breathed sulfate.
This process—the anaerobic oxidation of methane—would not be possible without such a close coupling between anaerobic methanotrophs and sulfate-reducing bacteria. Methane is a high-energy but very stable molecule: it is not easy to crack it open to release electrons and power metabolism. Anaerobic methanotrophs can do the job, but they end up releasing an overabundance of electrons as a result, leading to a backlog that would normally cause their metabolism to grind to a halt. One microbe's trash is another's treasure, though. The sulfate-reducing bacteria use the surfeit of electrons to turn sulfate into sulfide (which gives the sediment its putrid smell) and reap the energetic windfall that results. It is a classic symbiosis: the anaerobic methanotrophs enjoy a swift trash collection service, and the sulfate-reducing bacteria bask in an in-house power plant.
Our expedition to Hydrate Ridge showed that symbiotic methane consumption was happening not just in the sediment, where the phenomenon was first discovered, but also inside the carbonate rocks that form enormous mounds around methane seeps the world over. The interaction between the anaerobic methanotrophs and sulfate-reducing bacteria may take place on the microscale, but research in the Black Sea, the Gulf of Mexico and other locations has shown it is a pervasive process, soaking up roughly 80 percent of the methane emerging from the seafloor, building carbonate mounds on global scales.
The Origin of Teamwork
Earth's vast subsurface is rife with such examples of microbial interactions, and DNA sequences obtained over the past few years from microbial cells in groundwater and deep-sea sediments reveal just how interconnected these communities really are. As the number of DNA sequences has expanded, two startling conclusions have become increasingly inescapable. First, bacteria and archaea are far more diverse than anyone had imagined—the number of branches on the tree of life has exploded. But perhaps more surprising, their genomes are suspiciously small: many do not have enough information to build a fully functional cell or to complete the metabolic transformations that convert food into energy. “What we see all the time when we go into new environments,” says Laura Hug, a professor of environmental microbiology at the University of Waterloo, who was part of a team that discovered a number of previously unknown microbial species, “is that the entire community has the capacity for a certain function, like nitrogen cycling. All the pieces are there, but to identify a single organism that has all the pieces in its own genome—that's really unusual.”
The newly discovered cells' genomes often lack the ability to make all the amino acids needed to build their proteins or the nucleotides for constructing their DNA, suggesting they acquire these building blocks from neighboring cells with a surplus. These communities also appear to extract energy from the environment through a collective process: individual cells perform certain chemical conversions and pass the product down the chain to other cells for subsequent reactions. Sharing cellular building blocks and energetic resources in this way both requires and enables cohabitation among diverse organisms.
Although closely related cells still strive to acquire the same resources, the recent trove of genetic information suggests that at a larger scale, evolution has promoted specialization and collaboration. In much the same way that the global economy capitalizes on local strengths and the exchange of goods, groundwater and deep-sea microbial communities use division of labor to efficiently extract resources from sparse supplies, making harsh environments livable.
How do these vital collaborations arise in the first place? Some scientists believe that physical proximity within dense communities is a pivotal factor. As closely bound organisms reproduce, progeny remain nearby. With proximity comes the benefit of accessing parental resources, like a close-to-home college student taking advantage of laundry facilities. Natural selection favors the genes that make these shared resources when subsequent generations stay close to one another, but as physical proximity between parents and offspring decreases and genetically divergent cells enter the picture, freeloading mutants gain a selective advantage. They reap the benefits of shared resources without paying the costs of making them, taking over the community and lowering the overall rate of sharing. (This scenario is also called the tragedy of the commons, a term originally invented to describe a group of farmers with shared land; each individual farmer is motivated to have as large a herd as possible, leading to overgrazing and financial ruin for all.) These dynamics show that collaboration and sharing are favored when multiple generations of the same species remain closely co-located, a principle known as group selection.
But is this mechanism of group selection the ultimate explanation for the widespread microbial collaborations found in nature, or might other factors be at work? Clues have come from hundreds of feet above deep-sea methane seeps in the sunlit surface waters of the open ocean, where solar energy is abundant, but life-giving nutrients such as nitrogen and phosphorus are in short supply. Indeed, the tropical and subtropical surface waters were long thought to be ocean “deserts” until the late 1970s and 1980s, when scientists began to take a closer look at these environments and found teeming masses of microbes. Like the more recently detected groundwater and deep-sea sediment microbes, these surface ocean microbes have reduced genomes and cannot be cultivated without adding complex suites of nutrients to their growth media—telltale signs that these species need one another to survive. Yet whereas sedimentary microbes are stuck within dense cages of mineral particles—perfect conditions for group selection—microbes in the surface ocean float freely, constantly churned by their environment. Without reliable proximity to known neighbors, group selection cannot explain their cooperation. Some other force must be at work.
A Life-Changing Partnership
A single drop of water from the surface of the tropical ocean contains about a million microbes. One in 10 is likely a cyanobacterium known as Prochlorococcus, the smallest and most abundant photosynthetic organism on the planet. One of us (Braakman) has been peering into the DNA of Prochlorococcus, working with colleagues to understand how its metabolism has evolved over hundreds of millions of years. We created a metabolic family tree of this species by mapping variations in its metabolic network—the biochemical reactions that convert nutrient inputs to cellular building blocks—onto a genetic family tree that shows how the various kinds of Prochlorococcus are related. By comparing this merged metabolic family tree of Prochlorococcus subgroups with the large-scale gradients of light and nutrients where they are found, it became clear that evolution had selected for cells that harvested more solar energy and could best acquire sparse nutrients. At the same time, because more energy harvesting increases the throughput of carbon-based metabolism, cells became saturated with carbon. Energetically juicy molecules, packed with organic carbon, were released as waste—an exhaust valve on the powerful vacuum cleaner that could hoover up increasingly scarce nutrients. Prochlorococcus thus emerged as a cellular factory, soaking up sunlight and spitting out organic carbon waste.
This waste stream, in turn, became an attractive resource for microbes that cannot make their own food energy, including Pelagibacter, a distinct marine organism that, tellingly, is nearly as abundant as Prochlorococcus in the tropical and subtropical surface oceans. To investigate the relation between these two microbial groups, we also created a metabolic family tree for Pelagibacter and found an evolutionary path that completed the collaborative loop. Whereas Prochlorococcus consumes carbon dioxide and releases organic carbon compounds, Pelagibacter takes up those compounds and releases other molecules that Prochlorococcus can use for energy when the sun goes down. Both sides of this partnership recycle the waste of the other, extracting otherwise unused energy.
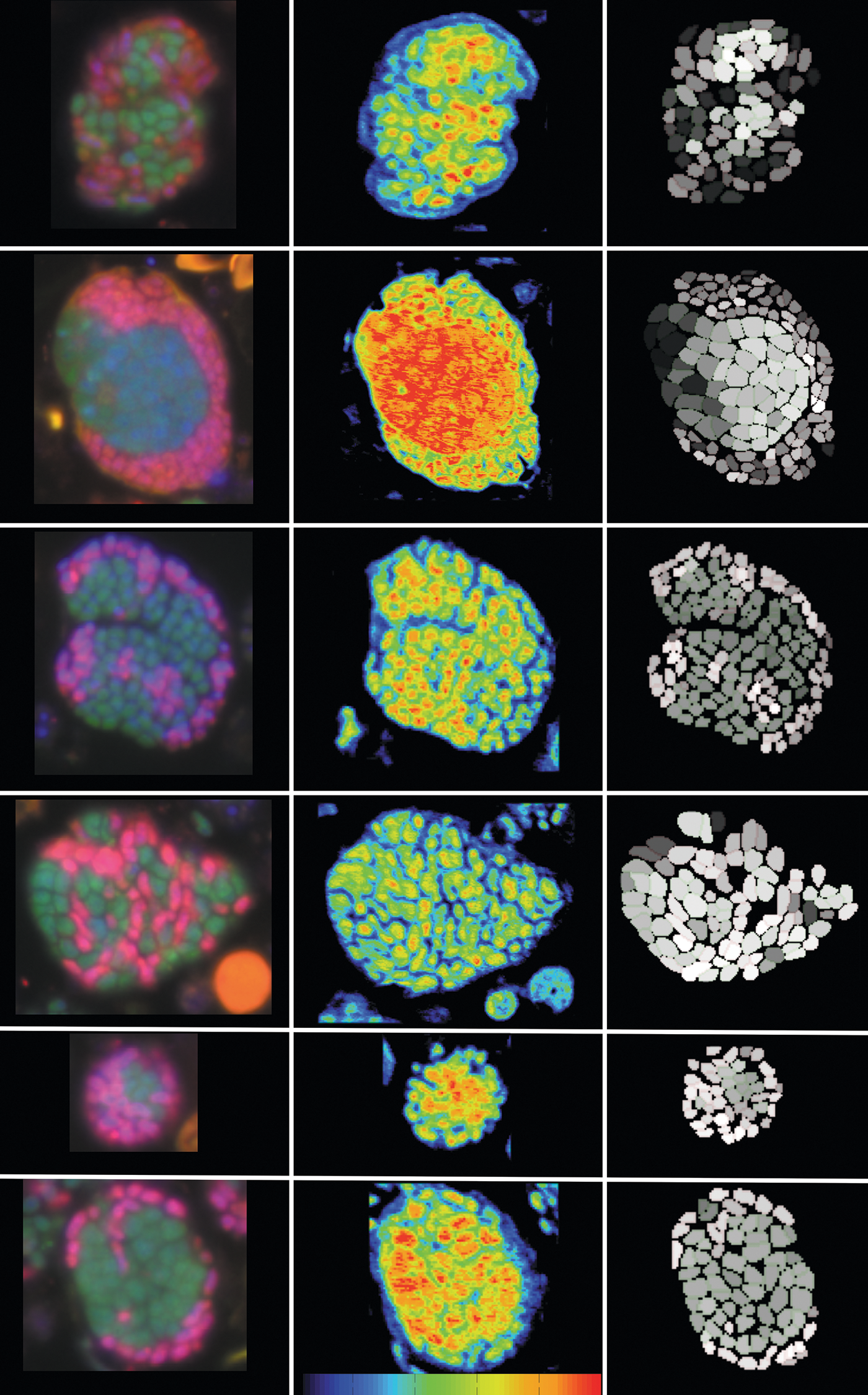
AGGREGATES of the anaerobic methanotrophs and sulfate-reducing bacteria that live in methane seeps are revealed using a variety of imaging techniques. From “Single Cell Activity Reveals Direct Electron Transfer in Methanotrophic Consortia,” by Shawn E. McGlynn et al., in Nature, Vol. 526; October 22, 2015
These findings, published in 2017, have important consequences for thinking about how microbial communities evolved in the surface oceans and other habitats. The implication is that as cells got better at collecting scarce nutrients, they drove the concentrations of those nutrients ever lower, dictating the terms on which all other organisms can use them. Freeloaders do not stand a chance, because cells that only consume but do not produce organic carbon are less proficient at acquiring other nutrients, such as nitrogen or phosphorus. Nutrient consumption and organic waste production are inextricably linked, strengthening the Prochlorococcus-Pelagibacter connection, which is bolstered by natural selection. This powerful arrangement shows that the evolutionary promotion of collaborative interactions does not apply only to tightly associated groups of closely related cells. At least in some cases, this selective drive may simply be a by-product—a self-amplifying feedback loop—of selection acting on individual cells.
The Prochlorococcus-Pelagibacter partnership may have emerged out of just a few small genetic changes, but its long-term effects were enormous. When the ancestors of Prochlorococcus and Pelagibacter colonized the oceans between 600 million and 800 million years ago, the waters were still largely devoid of oxygen and rich in iron. Iron is a requisite component of the photosynthetic proteins that ultimately generate oxygen, but it cannot dissolve and be wrangled into proteins when oxygen is around. This catch-22 would have kept photosynthetic organisms from expanding into the open ocean, where accessible iron would become scarce if they moved in and started making lots of oxygen. But Prochlorococcus's organic carbon waste products—fueled through growth alongside Pelagibacter—had a remarkable ability to bind iron, increasing its availability even in the presence of oxygen. Thus, we hypothesized that through the interplay between their organic waste and the critical iron, Prochlorococcus and Pelagibacter ultimately helped to pave the way for photosynthesis to oxygenate our planet's oceans. Life on Earth would never be the same.

Credit: Tami Tolpa
Ulterior Motives
Microbial interactions may not always be harmonious partnerships, however. Indeed, some scientists believe stable, mutually beneficial relationships may be the exception rather than the rule. “It's a dog-eat-dog world out there,” says biologist John McCutcheon of the University of Montana. “Even relationships that are temporarily beneficial in one context can lead to parasitism or competition in another, slightly different circumstance.” McCutcheon's Hobbesian worldview comes in part from the phenomenon he studies: endosymbiosis, or the wholesale incorporation of one organism into another. For example, the mitochondria that produce energy inside our cells were once free-living members of a group known as the alphaproteobacteria. Endosymbiosis has led to some of the most important innovations in life's history, generating the hallmark components of complex cells and paving the way for the evolution of plants and animals. Given these positive examples, “it's easy to imagine endosymbioses as a kumbaya kind of thing,” McCutcheon warns, “but I think it's a more exploitative interaction.” After all, he points out, evolutionary history is likely littered with failed attempts in which endosymbiosis trended toward either predation or parasitism.
Researchers have also found high rates of endosymbiont turnover, where, like a roommate that is just not working out, one incorporated species gets booted and a new one comes in, revealing an uneasy relation for both partners. McCutcheon's research amplifies the sense that interorganism interactions are indeed a dominant force while sounding a note of caution about their motives. “Every organism is looking out for itself,” he notes, “and not all interactions are good for everyone.”
There may also be a more fundamental downside to intricately connected microbial communities: if one member takes a hit, the rest of the network of mutually dependent microbes could be left vulnerable to collapse. In theory, metabolic linkages could render highly collaborative microbial communities more susceptible to failure than those made up of independent organisms that mind their own business.
Microbiologist Ashley Shade of Michigan State University and her colleagues examined 378 studies of soil, marine, freshwater, bioindustrial and animal gut microbiomes in an effort to develop general principles about community resistance to external disturbance and the ability to return to the baseline state. The researchers found that 56 percent of the investigations reported widespread metabolic changes after a disturbance—for example, exposure to heat prompted one soil-derived community of microbes to stop their usual consumption of nitrogen. Just 10 percent of these disrupted communities eventually resumed normal functioning. (These results should be interpreted with caution, however, because many of the compiled studies that looked at community resistance did not examine their eventual recovery. For those where recovery was examined, it is possible that researchers did not wait long enough to see things get back to normal.) Ultimately the biosphere is incredibly resilient and has always recovered from major disturbances—we would not be around otherwise—but much remains to be understood about how recoveries work, how quick they are and what long-lasting change persists.
We still have much to learn about the microbial communities that underlie the natural world and the role of collaborations. The results to date suggest that close metabolic partnerships drive evolutionary dynamics and open up vast new realms for colonization. But researchers have only just started looking at interactions beyond the microscopic scale, and placing these new findings into context in the real world continues to be a major challenge. How many species can interact in a meaningful way? How do the general principles shaping these interactions change in different environments or at different scales of space and time? A dense web of interacting microbes might mean that human-caused environmental influences could ripple through the entire network and lead to worldwide consequences we cannot yet anticipate. Continuing to decode these microbial networks is crucial as we enter an era of dramatic global change.
