

What is fear? The answer seems simple, yet a vigorous debate concerning its meaning has been playing out over the vista of affective neuroscience. This debate has a long history, but it was recently reignited by Joseph LeDoux, who proposed that we should not only redefine fear but also change the way we experimentally investigate this emotion. At the core of this debate lies the view that emotions are conscious, subjective states. For example, ‘feelings’ related to fear, such as horror or terror, are cognitively assembled conceptions of one’s situation, rather than preformed, innate mental states inherited from animals. LeDoux thus argues such complex states of the human brain cannot be studied in animals. Instead, he proposes that ‘defensive survival circuits’ that underlie defensive behaviors be the focus of research in animals. These hard-wired circuits are proposed to be orthogonal to subjective fear states that presumably involve higher-order circuits—they can modulate but do not determine the emotion. An equally provocative theory is Lisa Feldman Barrett’s ‘theory of constructed emotion’, which proposes that the human brain constructs instances of fear as a consequence of predicting and inferring the cause of incoming sensory inputs from the body (i.e., interoceptive and somatosensory inputs) and the world (i.e., exteroceptive inputs). Barrett proposes that a brain is continually projecting itself forward in time, predicting skeletomotor and visceromotor changes and inferring the sensory changes that will result from these motor actions. Probably most controversial about Barrett’s theory is that it proposes that fear, like other emotion categories, does not have a hard-wired neuroanatomical profile but is part of a dynamic system in which prediction signals are understood as ad hoc, abstract categories or concepts that are generatively assembled from past experiences that are similar to present conditions. In this view, the brain is a categorization machine, continually creating contextually relevant concepts that are appropriate to an animal’s niche.
These thought-provoking views seem to go against other prominent views, such as the basic (or primary) fear circuits theory of the late Jaak Panksepp and other celebrated luminaries in the field (for example, Michael Davis, Robert Bolles, O. Hobart Mowrer). For example, Ralph Adolphs emphasizes the universality of defensive behaviors, which adds credence to the view that fear circuits are mirrored across species and therefore partly innate. Michael Fanselow proposes that fear (and anxiety) can be placed along a threat-imminence continuum, which acts as a general organizing principle, and where threat intensity can be linked to motivational processes and defensive behaviors. Likewise, Kay Tye suggests that fear is a negative internal state that drives and coordinates defensive responses. These views see defensive behaviors as the manifestation of hard-wired fear (or survival) circuits and are controlled and modified by cognitively flexible circuits. While this debate has begun to wash up on the shoreline of clinical science and practice, there is still much needed agreement between the fields of basic and clinical science on how to define and investigate fear and anxiety. Here we asked some of the most influential contemporary scientists to discuss their perspective. Covering both human and animal research, each will present one argument for each of the discussion points below.
Q1: Dean Mobbs (moderator): How do you define fear and how is your definition supported by neuroscience?
On supporting science journalism
If you're enjoying this article, consider supporting our award-winning journalism by subscribing. By purchasing a subscription you are helping to ensure the future of impactful stories about the discoveries and ideas shaping our world today.
Ralph Adolphs (RA): Fear can only be defined based on observation of behavior in a natural environment, not neuroscience. In my view, fear is a psychological state with specific functional properties, conceptually distinct from conscious experience; it is a latent variable that provides a causal explanation of observed fear-related behaviors. Fear refers to a rough category of states with similar functions; science will likely revise this picture and show us that there are different kinds of fear (perhaps a dozen or so) that depend on different neural systems.
The functional properties that define the state of fear are those that, in the light of evolution, have made this state adaptive for coping with a particular class of threats to survival, such as predators. Fear has several functional properties—such as persistence, learning, scalability and generalizability—that distinguish emotion states from reflexes and fixed-action patterns, although the latter can of course also contribute to behavior.
The neural circuits that regulate an animal’s fear-related behavior exhibit many of these same functional properties, including in the mouse hypothalamus2, are initial evidence that this brain structure is not merely involved in translating emotion states into behaviors, but plays a role in the central emotion state itself. Neuropsychological dissociations of fear from other emotions show that fear is a distinct category.
Michael Fanselow (MF): Fear is a neural–behavior system that evolved to protect animals against environmental threats to what John Garcia called the external milieu (as opposed to the internal milieu), with predation being the principal driving force behind that evolution (for example, as opposed to a toxin). This is the organizing idea behind my definition of fear. The complete definition must also include the signals giving rise to fear (antecedents) and objectively observable behaviors (consequents). The neuroscientific support for this definition is that many signals of external threat, such as cues signaling possible pain, the presence of natural predators and odors of conspecifics that have recently experienced external threats, all activate overlapping circuits and induce a common set of behaviors (for example, freezing and analgesia in rodents). Equally important as neuroscientific support is support from fieldwork, which has repeatedly shown that behaviors such as freezing enhance survival in the face of predators.
Lisa Feldman Barrett (LFB): I hypothesize that every mental event, fear or otherwise, is constructed in an animal’s brain as a plan for assembling motor actions and the visceromotor actions that support them, as well as the expected sensory consequences of those actions. The latter constitute an animal’s experience of its surrounding niche (sights, sounds, smells, etc.), including the affective value of objects. Here ‘value’ is a way of describing a brain’s estimation of its body’s state (i.e., interoceptive and skeletomotor predictions) and how that state will change as the animal moves or encodes something new. The plan is an inference (or a set of inferences) that is constructed from learned or innate priors that are similar to the present conditions; they represent the brain’s best guess as to the causes of expected sensory inputs and what to do about them.
The function most frequently associated with fear is protection from threat. The corresponding definition of fear is an instance an animal’s brain constructs defensive actions for survival. A human brain might construct inferences that are similar to present conditions in terms of sensory or perceptual features, but the inferences can also be functional and therefore abstract, and thus they may or may not be initiated by events that are typically defined as fear stimuli and may or may not result in the behaviors that are typically defined as fear behaviors. For example, sometimes humans may laugh or fall asleep in the face of a threat. In this view, fear is not defined by the sensory specifics of an eliciting stimulus or by a specific physical action generated by the animal; rather, it is characterized in terms of a situated function or goal: a particular set of action and sensory consequences that are inferred, based on priors, to serve a particular function in a similar situation (for example, protection).
In cognitive science, a set of objects or events that are similar in some way to one another constitute a category, so constructing inferences can also be described as constructing categories. Another way to phrase my hypothesis, then, is that a brain is dynamically constructing categories as guesses about which motor actions to take, what their sensory consequences will be, and the causes of those actions and expected sensory inputs. A representation of a category is a concept, and so the hypothesis can also be phrased this way: a brain is dynamically constructing concepts as hypotheses about the causes of upcoming motor actions and their expected sensory consequences. The concepts or categories are constructed in a situation-by-situation manner, so they are called ad hoc concepts or categories. In this way, biological categories can be considered ad hoc conceptual categories.
Joseph LeDoux (JL): I have long maintained that conscious emotional experiences are, like all other conscious experiences, cognitively assembled by cortical circuits. Fear, for example, is a conscious awareness that you are in harm’s way. Activation of subcortical circuits controlling behavioral and physiological responses that occur at the same time can intensify the experience by providing inputs to the cognitive circuits, but they do not determine the content of the experience. The experience itself, in my model, is the result of pattern completion of one’s personal fear schema, which gives rise to some variant of what you have come to know as one of the many varieties subsumed under the concept of ‘fear’ that you have built up by accumulating experiences over the course of your life. Fear can even occur when some or all of the subcortically triggered consequences are absent: when the threat alone generates memory-based expectations that mentally simulate the missing elements, thereby pattern-completing your fear schema. Fear is often said to be universal. But instead what is universal is danger. The human expereince of being in danger is personal and unique. While other animals may have some kind of experience when in danger, it is not possible to scientifically measure what they experience, and if we could, it is unlikely it would be equivalent to the kind the of cognitively assembled personal awareness of being in harm’s way that humans experience. Such a cognitive account would seem necessary to explain, in one framework, the variety of threatening situations in which one can consciously experience fear (for example, predatory, conspecific, homeostatic, social, existential).
Kerry Ressler (KR): My definition of fear is one that is pragmatic and clinical, perhaps a ‘functionalist’ definition from Adolph’s perspective. ‘Fear’ is the combination of defensive responses—physiological, behavioral and (perhaps in the case of humans) the conscious experience and interpretations of these responses—that are stimulated by specific stimuli. In the case of experimental systems these stimuli are external cues, but presumably in humans can have internal representations as well (thoughts and memories that can be fear-inducing cues themselves). Such fear-inducing cues result in active defensive responses that gradually subside when the stimulus is no longer present. Clinically, fear can be thought of as mirroring the response to a specific cue (for example, the fear of snakes), while anxiety is a more long-lasting phenomenon that may not be specific to overt cues. Decades of preclinical neuroscience studies examining mechanisms of Pavlovian fear or threat conditioning have, in conjunction with human neuroimaging work, indicated the involvement of multiple brain regions in communication with the amygdala and its downstream connections in support of the ‘hardwired’ regulation of subcortical and brainstem areas mediating the cardiovascular, respiratory, autonomic nervous system, hormonal, startle, freezing and other behavioral ‘fear’ or ‘threat’ reflexes.

Credit: Ekaterina Nosenko Getty Images
Kay Tye (KT): Fear is an intensely negative internal state. It conducts orchestration of coordinated functions serving to arouse our peak performance for avoidance, escape or confrontation. Fear resembles a dictator that makes all other brain processes (from cognition to breathing) its slave. Fear can be innate or learned. Innate fear can be expressed in response to environmental stimuli without prior experience, such as that of snakes and spiders in humans and to predator odor in rodents. Fear associations—primarily studied in the context of Pavlovian fear conditioning—are the most rapidly learned (one trial), robustly encoded and retrieved, and prone to activate multiple memory systems. Given its critical importance in survival and its authoritarian command over the rest of the brain, fear should be one of the most extensively studied topics in neuroscience, though it trails behind investigation of sensory and motor processes due to its subjective nature. Watching others exhibit the behavioral expressions and responses of fear may invoke emotional contagion or support learning about the environment. The usage of the term ‘fear’ in the field of behavioral neuroscience has taken on a related—but distinct—meaning through the extensive use and study of a very stereotyped behavioral paradigm originally termed ‘fear conditioning’. Fear conditioning is arguably the most commonly used behavioral paradigm in neuroscience and has been most comprehensively mined in terms of neural circuit dissection with rodent models but has also been used in humans, primates and even invertebrates. Fear conditioning refers to the Pavlovian pairing of a conditioned stimulus (most often an auditory pure tone) with a foot shock that is most often presented upon the termination of the conditioned stimulus.
Q2: How does your theory of fear separate neural circuits for feeling, perception and action?
RA: I don’t claim to have a theory, but in my view fear, feeling, perception and action are all distinct. Fear causally interacts with many other processes, including perception, action planning, attention, memory and others. But it is distinct in that we can manipulate fear independently of many other cognitive variables. Losing perception, as in blindness, doesn’t make you lose fear, merely the ability to induce it visually; losing all behavior, as when paralyzed, also doesn’t make you lose fear; similarly for memory and other processes. It is important to note that a state of fear by itself does nothing: it needs to connect with all these other processes to result in behavior (as is the case for perception, attention, etc., themselves). Most important is the distinction between feeling fear (the conscious experience of fear) and the functional state of fear (the state that explains all the effects a threatening stimulus has on cognition and behavior). I’m agnostic about how these are related, but I think for methodological reasons, for example, the ability to study fear in nonhuman animals, we need to keep them conceptually separate. It is also very difficult to distinguish the neural correlates of feeling fear and the functional state of fear. All of the above suggest some cognitive architecture defined by constitutive and causal relations between processes. How this is actually neurally implemented in no doubt varies between phyla and classes; fear in an octopus will have very different neural details than fear in a human or a rat.
MF: It doesn’t. The relevant circuit integrates them; perception of threat leads to feelings and to actions. Activation of the fear state also feeds back on perceptual systems, altering how they react to environmental stimuli. The perception of threat is a critical determinant of both the magnitude of fear and the topography of defensive behavior. Note that not all actions stem from feelings, but all fear-related feelings lead to some change in action. If they didn’t, they would lose biological meaning and, to the extent that feelings require energy, they would be eliminated by evolution. A complete circuit connects and integrates these components into effective defensive patterns.
LFB: In my view, this is not the optimal question to ask about fear because it rests on an unfounded assumption that the brain is best understood as collections of neurons, grouped together in anatomically separate systems (neural circuits) for perceptions, mental events, feelings and various types of action (for example, freezing, running, etc.), which pass information back and forth to one another like a baton in a relay race. My research approach is guided by the alternative assumption that the brain should be understood as a complex dynamical system that is composed of elements: circuits or subnetworks made of neurons and supporting glial cells. These elements do not function independently of one another, because their arrangement and organization change dynamically. Even the neurons that constitute change dynamically. The brain, as a dynamical system, is continuously traversing through a succession of events, referred to as its state space, which is specified as values for a set of features that describe the system’s current state. Features are physical (for example, neural, physiological, chemical) and mental (perceptual, affective, cognitive, etc.). In this view, the brain works by prediction and correction rather than through stimulus and response. Within the dynamics of a particular state of the system perceptions are the result of motor preparation, rather than the other way around (as suggested by a stimulus–response approach).
JL: In my scheme, fear is the feeling of being afraid. I would refer to ‘perception’ and ‘action’ in this context as ‘threat detection’ and ‘defensive responding’. I view the experience of fear and behavioral reactions as separate consequences of threat detection and mediated by different but interacting circuits. Threat detection obviously starts with sensory processing, research on which is informative in illustrating the relationship between stimulus processing, behavior and experience. For example, studies of visual perception in patients with blindsight show that the path to conscious perceptual experience can be dissociated from the path to behavior. This suggests that the correlation of perceptual experience with behavior in healthy brains may be due to parallel processing of sensory information by different systems and does not necessarily mean that the experience and behavior are entwined in the brain. Perceptual researchers thus tend to be cautious when extrapolating from behavioral responses to experience. In terms of fear, blindsight is again informative. These patients respond to threats but do not report awareness of the threat stimulus or conscious feelings of fear; self-report of conscious feelings in such patients correlates with neocortical activity. Similarly, in subliminal-stimulation studies of healthy humans, threats activate subcortical defensive circuits involving the amygdala and elicit physiological responses in the absence of stimulus awareness; feelings are not reported even when specifically asked about. The circuits that control behaviors that are only sometimes correlated with fear experiences are thus not necessarily the circuits that underlie the experiences. When we label these circuits and behaviors with the term ’fear’ we propagate conceptual confusion.
KR: I think that we can, at a neuroscience level, make some distinctions between the sensory components (for example, sensory thalamus and cortex: feeling), integrative cognitive components (for example, associative cortex and medial prefrontal cortex: perception) and reflexive and behavioral components (for example, amygdala, striatum, brainstem: action). However, how these distinct circuits map upon conscious vs. behavioral aspects of fear processing may be more difficult to parse. Progress in dissecting the neural connections of fear and threat has contributed to our understanding of how they regulate the autonomic, physiological and behavioral activity patterns that together comprise the ‘fear reflex’, which appears to be highly conserved across species. Some aspects of these different components are clearly represented in similar areas—for example, medial prefrontal cortex and amygdala activation are seen with threat perception in humans, but are also clearly involved in actions underlying threat behaviors across species—whereas other regions, for example, brainstem nuclei, may be involved primarily in the action component of the fear process.
KT: Initial information flow arrives via sensory inputs that propagate to limbic circuits (for example, amygdala), which then feeds forward to downstream targets (for example, striatum, basal ganglia), where emotional state combines with threat imminence to promote action selection. Limbic signals can then feed back onto the sensory systems to alter perception. Fear itself does not map onto an individual motor output; it is an intermediate process that links sensory processing to action selection. My current conceptual model consists of three psychological processes that determine importance (or salience), valence and action, respectively. These three processes are mediated by different circuits. For example, if a grazing deer hears a twig snap, it must initially assess the importance of the stimulus. If it is in a clear landscape with nowhere for a predator to hide, then the stimulus may be deemed unimportant and the deer may go on grazing. If the deer sees a familiar conspecific, then it may interpret the stimulus as a positive valence signal, prompting selection of agonistic social behavior or approach. If there is dense brush, then the potential threat of a predator signaled by the stimulus may trigger an internal state of fear. Given a fear state, the outcome depends heavily on threat imminence. For example, if the predator is far away or its location is unknown, it may be most adaptive to hide or freeze to avoid detection by the predator. If the predator is at an intermediate distance where detection is likely or has already happened, then escape may be the best strategy. If the predator is mounting an attack, then defensive behavior to fight off the predator may be the best response.
.jpg?w=2000)
Credit: Getty Images
Q3: Are there different defensive circuits (for example, predatory vs. social, survival circuits, reactive vs cognitive fears), and if so, are they orthogonal or synergistic? What is the evidence for your position?
RA: Yes, I think there is very good evidence that there are neural circuits specialized for subtypes of fear. Fear is not one thing. For instance, a circuit involving the superior colliculus and periaqueductal gray has been dissected in some detail for mediating fear behaviors elicited by the sight of aerial predators in rodents. Conversely, the ventromedial hypothalamus has cell populations that participate in states of fear and respond to sounds or odors of conspecifics but not to aerial predators. There are also different circuits relating to threat imminence (anxiety, fear, panic). Work in humans with amygdala lesions has dissociated fear of teloreceptive stimuli (snakes, spiders, etc.) from fear of interoceptive stimuli (suffocation). To the extent that different types of threat require different adaptive behaviors, they would constitute different functional states—and this functional specialization should be reflected in the neural circuits. These relatively ‘dedicated’ neural circuits for subtypes of fear are subcortical, whereas cortical involvement is likely to feature ‘mixed selectivity,’ such that the same cortical neurons can encode the multiple actions that might need to be taken in an adaptive response to fear, depending on the circumstances.
MF: Yes. For example, the taste aversion–disgust–toxin avoidance system (Garcia’s internal milieu defense) is distinct from predatory defense (external milieu). In a nice demonstration of this, Bernstein’s lab showed that within the basolateral amygdala, taste (conditioned stimuli) and toxin (unconditioned stimuli) converge on different sets of neurons than contextual conditioned stimuli and shock unconditioned stimuli. This illustrates the common error of considering the basolateral amygdala as isomorphic with ‘fear’. It is not; it mediates several aversive and appetitive motivational systems that involve different cells and microcircuits within the amygdala. Another concern about purely amygdalocentric views is that not all antipredator defensive modules are equally dependent on the amygdala. For example, I proposed a circa-strike–panic defensive module that depends more on periaqueductal gray than amygdala. This model anticipated the finding that CO2-induced panic occurs in a patient with bilateral loss of the amygdala who otherwise is severely deficient in fear reactions. Interactions between different aversive systems, much like interactions between appetitive and aversive systems, are often inhibitory because the systems serve different functions and one function may need to take precedence over another; for example, inhibition of the pain or recuperative system via analgesic circuitry is part of the fear and defense system. But there is also convergence. In rodents, defense against predators (interspecies) and alpha males (conspecifics) activates very similar brain structures and behaviors, suggesting that there was substantial convergent evolution of these defenses. One reason my essay (Supplementary Information) provides for a rich (six-part) definition of fear is to help distinguish fear from other systems.
LFB: Neuroscience research on motor control has revealed that motor actions are not triggered by simple, dedicated circuits, but are assembled within a flexible neural hierarchy whose motor modules are in the spinal cord. I hypothesize that the same may be true for visceromotor actions. In this view, attempts to build taxonomies of simple defensive circuits are not scientifically generative. The presence of flexible neural hierarchies means that each behavior—such as freezing, fleeing and fighting—is not the result of one specific circuit, but instead may be implemented in multiple ways. In my view, a brain, as a single dynamical system, has the core task of regulating skeletomotor actions as well as visceromotor actions within the body’s internal milieu that supports those actions. This idea suggests that there are degenerate assemblies for each behavior, even in the same situation. Furthermore, the neurons that process sensory inputs (for example, in V1, primary interoceptive cortex) and the neurons that represent affective value all function in the service of actions and carry information about those actions, and therefore are part of the flexible hierarchy for action control.
JL: Nathaniel Daw and I recently proposed taxonomy of defensive behaviors and their neural underpinnings that might provide an organizational framework for considering some of the diverse levels of analysis implied in the present question. Included are reflexes, fixed reactions, habits, action–outcome behaviors and behaviors controlled by non-conscious and by conscious deliberation. For example, species-typical responses to predatory and social cues can be thought of as fixed reactions that are ‘released’ when different, but to some extent overlapping, subcortical ‘survival circuits’ are engaged. Also relevant are circuits that signal challenges to survival monitor homeostatic imbalances and initiate restorative behaviors. Instrumental, habitual behaviors are fixed but have to be learned and involve corticostriatal circuits, whereas action–outcome instrumental behaviors are learned but flexible and use different corticostriatal circuits. Deliberative instrumental responses are prospective and model-based, and they engage prefrontal circuits; here, non-conscious deliberation about danger allows rapid mental simulation of possible solutions, whereas in slower conscious deliberation, the experience of fear can guide future planning and action.
KR: For brevity, I will focus on ‘the’ amygdala, which is actually a complex of several cell clusters (nuclei) and is conserved from the most primitive mammals and in most vertebrates. It receives neural projections from essentially all sensory areas of the brain, as well as from memory-processing areas in addition to association and cognitive brain regions. It sends projections back to many of these areas, but most interestingly, also communicates with an array of brainstem and other subcortical areas. Notably, all of these circuits are involved in both defensive and appetitive behaviors, not to mention predatory vs. social behaviors, etc. Recent fascinating work has shown that even within the same subregion of the amygdala, neighboring cells can have opposing functions or more-nuanced functional differences; for example, they may respond preferably to proximal vs. distal threats. Such findings suggest that parallel information pathways, for example different cells encoding ‘fear-on’ vs. ‘fear-off’ information, flow through basolateral and central amygdala nuclei. Furthermore, the same cells that ‘turn off’ a fear response may be responsible for activating positive emotions, such as appetitive or even addictive behavior. Thus, these information channels may be better appreciated as underlying approach vs. avoidance related behaviors and drives. However, it is also possible that as such behaviors are parsed at a neural circuit level, they won’t match well onto our historic terminology of defensive circuits as outlined.
KT: Synergistic. Everything is connected in the limbic system, if not through direct reciprocal connections, then through neuromodulatory systems. Circuits that mediate different types of fear are likely to converge onto some common pathways, before diverging again for action selection. For example, animals can learn to fear an environmental stimulus through firsthand experience but also through observing others. We know that the basolateral amygdala (BLA) is a critical nucleus for translating sensory information into motivational significance for associations learned through direct experience and that observational fear learning requires both the BLA and the anterior cingulate cortex. The anterior cingulate cortex’s role is to interpret the demonstrator’s distress and send this signal to the BLA, where associative learning takes place.
Q4: How does (or can) your perspective fit with the others’ perspectives?
RA: My functional emphasis is probably closest to the views of Mobbs and Fanselow. I particularly like threat imminence theory, which is of course a functional theory. My view of fear as a state that is distinct from the conscious experience of fear seems aligned with LeDoux’s view with respect to that emphasis. This is a bit ironic, since I disagree with LeDoux’ conclusions (he redefines ‘fear’ to mean ‘the conscious experience of fear’), but I think he has written most clearly about the distinction, which is important. I would actually reinterpret his view as being about how we recognize that an organism is in a state of fear. We recognize this state in ourselves by having a conscious experience of fear; we recognize it in other people from their verbal reports or behavior; and we recognize it in animals from their behavior. If we want to be consistent, we should apply whatever meaning of ‘fear’ to both other humans and to animals, since the evidence is of the same type. Ressler’s and Tye’s views stay closer to the neurobiology, and I certainly share the view that a lot of questions about fear are empirical matters, mostly still needing resolution. There is no question that the science of fear, even in the absence of any agreement on conceptual or theoretical issues, will make progress and indeed will inform the conceptual and theoretical issues. I would agree that it’s productive to just get on with the neuroscience even without agreement about the philosophical issues; but I also think we need to continue to take stock and discuss the philosophical issues to get a sense of where we’re heading. Feldman Barrett’s view both shares some strong agreement with mine and is completely opposed. I share her emphasis on the context-dependency of emotions and, in particular, her attack on the notion that we can ‘read out’ emotions from facial expressions (indeed, we just co-authored a paper on this). But I disagree with her notion that there are no objective criteria to decide whether an animal or person is in an emotion state or in a particular type of emotion state.
MF: Like Adolphs’ approach, my approach emphasizing evolutionary demands is a take on functionalism; indeed, my first paper on predatory imminence was titled, “A functional behavioristic approach to aversively motivated behavior.” I resonate completely with Adolphs’ sentiment that “emotions are states of an organism that are defined by what they do.” I note that both Adolphs and LeDoux are critical of behavioristic approaches, but their criticism is leveled at radical behaviorism. My behaviorism is a product of Tolman’s cognitive behaviorism that emphasized purpose in behavior, although Tolman was more focused on immediate or proximal function (how do I get food here) as opposed to ultimate function (why do I seek food). Indeed, fear-related actions were phylogenetically programmed because they had a high probability of success over many generations, but the actions may be maladaptive in an immediate situation. This also means that any individual instance of these programmed behaviors may not be effective in the current situation. That is why any particular instance of fear behavior may seem, and actually be, irrational in the present moment. My approach appears to be in direct contradiction with both Feldman Barrett and LeDoux’s ideas that fear is entirely a higher-order conscious construction. The adaptive function of consciousness is typically viewed as providing flexibility and supporting deliberative, proximally rational, behavior. I think this stands at odds with the necessary features of life in the face of threat. Reactions have to be immediate; any time spent in deliberation increases the likelihood of death. Therefore, these fear reactions are phylogenetically programmed responses. When faced with a predator, there is no time to acquire behaviors based on trial and error and no time for novel planning. The contrast with Tolman is again instructive. Tolman emphasized variable means to fixed ends; if you have a cognitive map that reveals the location of food, the animal may use many different ways of getting to that food. The idea is quite similar to Feldman Barrett’s description of one-to-many mapping in motor systems. But Tolman’s theory was based on empirical work with a food reinforcer, where considerable flexibility is not only tolerated but beneficial: you don’t die if you miss one meal, and trying out something new may lead to a richer patch or a nutrient unavailable in the preceding meal. The demands of defense are entirely different. Hence, the rodent’s most studied food-getting response, lever pressing, is virtually impossible to investigate in the frightened rat.
LFB: Empirically, the scientific findings constitute a small subset of what remains to be discovered about the neurobiological basis of fear. My scientific approach differs substantially in its guiding ontological commitments than those that guide current research on the nature of fear.
JL: Each of the participants has laid out a cogent argument for their position. I enjoyed reading the essays, and I learned something new about what each author thinks. My ideas about the conscious experience of fear overlaps with Barrett's, as we both view fear as a cognitively assembled state that is based on mental models and conceptualizations of situations. For me, the other factors or ingredients that contribute to fear, such as brain arousal and feedback from body responses, modulate but do not determine the quality of the experience. On the other hand, my ideas about the role of brain areas such as the amygdala in detecting threats and initiating body reactions, and on the role of resulting motivational states that guide instrumental actions, are largely compatible with the views of the other contributors. Much of what we disagree about is semantic—in the presence of a threat, is fear the experience itself or all of the various consequences triggered by the threat? But to say the differences are semantic does not mean they are unimportant. Words are powerful. They underlie our conceptions and shape the implications of our theoretical points of view, and they influence what others conclude about our research. We should do our best to eliminate ambiguity and confusion in our scientific word choice. Our lexicon provides us ways to do this, and we should make use of the subtlety of our language when we use it scientifically. An easy way to start is to avoid using mental state terms to describe behaviors that are not based on mental states. In humans we can make these distinctions, and should then should certainly avoid using mental state terms to describe behavior in animals when in humans similar responses are not controlled by subjectively experienced mental states. I believe that words like threatening stimuli, defensive responses and defensive survival circuit characterize stimulus-response relations in animals better than fear stimuli, fear circuits and fear responses.
KR: In most ways, I agree with the other perspectives, in that I feel everyone is stating similar aspects of a broader shared understanding, but with nuanced differences. I think my perspective is most focused on the observation that in human neuropsychiatry research, the science of aversive behavior and fear-related disorders, along perhaps with appetitive behavior and addiction, is the most mature for clinical translation. Specifically, I agree with Adolphs’ idea that a “functionalist view of emotions like fear requires an interdisciplinary approach.” I agree with Fanselow’s defining characteristics of fear—a formalistic approach which I believe has much utility, in particular with regard to the differential experiential states that distinguish different functional modes between anxiety, fear and panic. I agree with Barrett that the features of fear “include some set of physical changes (autonomic nervous system changes, chemical changes, actions, etc.) and sensations that become perceptions of the surrounding world and the body.” I agree with LeDoux that “fear is a conscious experience in which you come to believe that you are about to be harmed” and with Tye on the importance of a conceptual model consisting of “three psychological processes that determine importance (or salience), valence and action, respectively.” While I also agree with many of the nuanced, philosophical, psychological, behavioral and neuroscience-based definitions, I don’t want to lose sight of how much progress has been made and how powerful the concept of ‘fear’ is to translational neuropsychiatry.
Q5: Do current behavioral assays for the study of fear restrict our ability to improve our understanding of fear?
RA: The contemporary assays are seriously flawed in that they compare apples and oranges between studies in animals and studies in humans. There are quite a number of behavioral assays for fear in animals, essentially none of which are used in studies in human studies, which instead typically use verbal reports as the ground truth. Since it’s impossible to use verbal reports in animals, the solution seems in principle straightforward: we need to adapt the behavioral batteries from animal studies to studies in humans. Only a few studies have attempted this. An additional challenge of course is ecological validity. Mobbs’ study of moving a tarantula closer and closer to your foot while you are in the scanner is a rare but classic success in this direction. The problem also extends to the stimuli used. There are many studies that present human subjects with facial expressions of emotions or that have them read short vignettes. Those studies may show something about social perception or people’s semantic knowledge about the concept of fear, but they do not assess the actual state of fear. I am quite concerned about the inadequacy of most experimental protocols to study human fear, which have disconnected the study of fear in humans from the study of fear in animals. Human studies need more ecologically valid stimuli and better behavioral assays, in particular ones that do not rely on verbal report and that can be argued to have some homology to the behavioral assays used in animal studies.
MF: Pavlovian fear conditioning is a natural component of how prey recognize predators and it works great in the lab. But its success comes with dangers. One of these dangers is that it has led to disproportionate emphasis on one module in the threat continuum (post-encounter–fear) and our knowledge of the other components (circa-strike–panic and pre-encounter–anxiety) lags behind. Perhaps an even greater danger is the tendency to treat procedure as isomorphic with process. Procedurally, fear conditioning is defined as pairing a neutral stimulus with an aversive one, but this procedure will not invariably condition a fear state because not all aversive stimuli support engagement of the antipredator defensive system. A toxin is clearly an aversive stimulus, but pairing a neutral flavor with a toxin leads to palatability shifts that reduce consumption and not an antipredator defense. Likewise, some shocks are sufficiently novel and powerful to condition fear but others are not; a mild shock may well be annoying but insufficient to condition fear. A rat’s behavior is more flexible with a very weak shock, but that flexibility is progressively lost as shock intensity increases. I take this loss of behavioral flexibility as diagnostic of a fear state. Therefore, one must be cautious when choosing shock intensity or letting subjects choose shock intensity. Additionally, other commonly used outcomes in human fear studies, such as loss of money, are unlikely to tap into the neural systems that support antipredator defense.
LFB: Contemporary paradigms, guided by the notion of simple, dedicated neural circuits for fear arranged in a single taxonomy, restrict the study of fear in several important ways. First, instances of fear are typically studied in laboratory settings that differ strongly from the ethological contexts in which they naturally emerge. All potential actions have an energy cost, and an animal’s brain weighs these against potential rewards and revenues in a particular context. Economic choices about actions, therefore, are necessarily influenced by a number of situation-specific considerations about an animal’s state and the state of the environment, most of which are held constant in the typical laboratory experiment. These factors not only influence which defensive action is executed (as suggested by some taxonomies of defensive behaviors), but also how any given action is implemented. Ignoring these factors make the neural causes of defensive actions seem more atomistic than they actually are, and as a consequence, most contemporary paradigms are insufficiently holistic (see my answer to Question 2). Second, contemporary paradigms confound things that should be kept separate. For example, it’s important to distinguish affect and emotion. Affective features such as valence and arousal are best thought of as low-dimensional summaries of higher-dimensional interoceptions that result from allostasis; valence and/or arousal might be intense during episodes of emotion but are not specific to those episodes. Because allostasis and interoception are continually ongoing in an animal’s life, valence and arousal are mental features that may describe every waking moment of that life. For this statement to make sense when comparing human and non-human animals, it is necessary to distinguish a brain’s capacity for consciousness (an experience) and its capacity for awareness (the ability to report or reflect on an experience); relatedly, it is important to distinguish perceiving the sensory features of the immediate context in a particular way from being aware of that perception (for example, an awareness of perceiving threat) and from the awareness of being frightened. It’s also important not to confound a threatening stimulus with the context in which the threat emerges, as often occurs in taxonomies of fear; brains don’t perceive stimuli, they perceive sensory arrays, i.e., ‘stimuli’ in context. And perhaps most importantly, one should not confuse observation and inference. Scientists measure things like skeletomotor actions (such as freezing) and the visceromotor actions that support those skeletomotor actions (such as changes in heart rate), which they might refer to as ‘fear’; correspondingly, they measure the change in neural firing that supports those actions, which they might refer to as ‘fear circuitry’. This approach confounds what is observed (for example, freezing, changes in heart rate) with their inferred cause (for example, fear). The science of fear would be more productive and more generative if the two were not routinely confused. When a scientist observes actions and infers an instance of fear, the scientist is engaging in emotion perception. Fear is always a perception—an inference—whether on the part of a scientist observing an animal’s actions, a human observing another human’s actions, or an animal making sense of its sensory surroundings as part of action control. No changes in the autonomic nervous system or skeletomotor actions are, in and of themselves, meaningful as fear. A brain makes them meaningful as fear with inferences (which can also be described as prediction signals or ad hoc concepts). An animal’s brain—human or otherwise—makes these inferences without awareness of doing so. From this perspective, understanding the neurobiological basis of inference is part of understanding the neurobiology of fear.
JL: A staple of research on fear has, of course, been the ‘fear’ conditioning paradigm. It has generated a large amount of useful information about how the brain detects and responds to danger. It can also be used to probe human participants about conscious experiences. But in studies of non-human animals, for reasons discussed in detail elsewhere, researchers can only measure behavioral and physiological responses. Because similar responses, including amygdala activation, can be elicited in humans with subliminal stimuli that are not consciously perceived and that do not engender reports of fearful feelings, the experience of fear would not seem to be driving the responses. For this reason, the amygdala circuit might be better thought of as a threat circuit or defense circuit than a fear circuit. Thus, the limits lie not in our paradigms; rather, the paradigm exposes the limits of what can be learned from animals versus humans when using these paradigms. Our understanding of fear is, however, limited by other things. One is the fact that truly frightening and traumatizing situations, for ethical reasons, cannot be used in laboratory studies of fear; milder proxies only give us hints, as brain responses do not scale linearly with stimulus intensity. Another is conceptual complacency and loose use of language. As noted above, popular views of fear and fear conditioning are tethered to Mower and Miller’s conceptualization dating back to the 1940s. The term ‘fear conditioning’ implies that the task reveals how fear arises. If one thinks of fear as a conscious experience, as I do, fear conditioning (or what I call ‘threat conditioning’) can in principle be used in animal studies to help understand processes that contribute indirectly to fear; but it cannot reveal the mechanisms underlying human fearful experiences, which can only be studied in humans (I do not deny animal consciousness as a natural phenomenon but question whether we can study this scientifically). I believe that the use of mental-state words like ‘fear’ to characterize behavioral control systems inevitably creates confusion and leads to misplaced expectations about what animal research can and cannot tell us. Thus, if someone uses the word ‘fear’, then he or she should clarify the intended meaning of ‘fear’ each time the term is used (for example, adding adjectives such as ‘conscious’ or ‘non-conscious’ or ‘explicit’ or ‘implicit’) to avoid confusion. Separating conscious ‘fear’ from non-conscious ‘threat processing’ from the start would avoid such confusion.
KR: The most common current approaches to study fear in preclinical model systems are based upon Pavlovian fear conditioning models—examining the different memory-related constructs of acquisition, expression, extinction, etc. of a fear memory—and use behavioral metrics of freezing, avoidance and startle. Similarly, in most human models, laboratories have sought to perform controlled experiments but generally using self-report or physiological outcome measures (for example, electrodermal skin response, heart rate or acoustic startle). A limitation to most translational studies is that the human and model-system studies generally do not use the same paradigms and same outcome metrics. Furthermore, using well-controlled learning paradigms makes it harder to explicitly define pathways and agreed-upon circuits related to innate or unconditioned fear cues, processes and behaviors, particularly in animal model systems. Generally, the more controlled and reductionist the experimental paradigm, the harder it is to observe and quantify natural threat response patterns and their underlying biology.
KT: I think having a very stereotyped behavioral paradigm for Pavlovian fear conditioning has facilitated reproducibility and a deeper dive into the anatomy and mechanism (for pairing pure tones to co-terminating foot shock in rodents). However, there are many other types of fear that have been understudied or not yet studied at all, leaving us with more depth and less breadth in our understanding of fear. At this point, the vast majority of publications on ‘fear’ refer to a very specific paradigm that is only a tiny subset of the neural mechanisms of this emotional state.
Q6: Can animal models inform us about human models of fear (and vice versa)?
RA: I would say studies in animals are essential to understanding fear, since they allow much better measurements and manipulations than is the case in humans—neither are ‘models’ of anything. The animal studies investigate animal fear; the human studies investigate human fear. No doubt there will be both similarities and differences between any different species, and some animals will have functionally defined fear states that are completely absent in others (animals that don’t live in an environment with aerial predators will not have the circuit involving the superior colliculus that processes that type of threat in mice). The reason I actually favor animal studies over human studies is that they can simplify what we are looking for. As I noted earlier, studies in humans typically mix the study of fear with the study of the concept of fear, the conscious experience of fear, or the verbal report of fear. A mouse certainly doesn’t have the verbal report, is unlikely to have the concept, and we don’t know how to measure its conscious experience—when confronted with a threat, it is just in a functionally specified state of fear. It is also much easier to induce ecologically valid emotions in animals (they don’t know they are in an experiment), and it is much more difficult for animals to volitionally regulate their emotions. For all these reasons, studying genuine, intense emotions in animals is far easier than studying them in humans and should be the place where neuroscientists start.
MF: Absolutely and they have. Wolpe’s development of exposure-type therapy was drawn from animal work by Pavlov and Hull and still stands as the signature treatment for anxiety disorders. Mobbs has provided a sophisticated expansion of predatory imminence theory that allows it to capture many of the unique features of human emotion.
LFB: Animal models can inform us about human instances of fear, but currently there are several obstacles. First, most animal studies are performed in just a few model species and fail to consider the similarities and differences in brain-based and niche-based features of different species and as model systems for neurotypical human brain development and function. The computational role of most major brain parts remains conserved across the vertebrate lineage, and all brains can be described as automatically and effortlessly forming inferences (i.e., ad hoc concepts) to categorize anticipated sensory inputs and guide action. But species may differ in the type of concepts that a brain can construct, due to general brain-scaling functions and the information available in an animal’s niche. For example, the human brain has expanded association cortices compared to other primates, enabling increased information compression and dimensionality reduction; this suggests that human brains may be able to create multimodal summaries characterized by more abstractio. This hypothesis in no way diminishes the importance of survival-related behaviors in human emotion, nor does it invalidate the importance of studying survival-related behaviors in animal models for the purposes of understanding the biology of human emotion. It does suggest, however, that solving the puzzle of human emotion—and human evolution more generally—may require a science of ‘emotion ecology’ that attempts to understand species-general and species-specific processes. Moreover, experimental animals are typically reared in impoverished laboratory settings with fewer opportunities to encounter the range of sensorimotor challenges than are typical in natural ethological contexts; this likely impacts brain wiring during development, prompting the question of whether lab animals are even ‘neurotypical’.
JL: The answer to this question is obviously yes, but the details depend on the animal in question and what one means by fear. Invertebrates can potentially inform us about cellular and molecular mechanisms of threat learning in mammals, including humans. Non-primate mammals can potentially inform us about circuits that detect threats and control various responses (for example, reactions, habits, instrumental actions). Non-human primates can potentially inform us about cortical circuits that underlie deliberative cognition. But in each case it is important to verify, to the extent possible, the relevance of the findings to humans by doing studies that approximate the animal studies in humans, albeit with less neurobiological detail. Human research is also necessary to study the conscious experience of fear and other emotions. This is true for at least two reasons. First, methodological barriers limit the assessment of consciousness in non-human animals. We can, as Jeffrey Grey put it, only creep up on consciousness using behavioral proxies in non-humans. Flawed though it is, verbal report is a powerful tool in humans. We can typically respond verbally or non-verbally to information which we are conscious of, but can only respond non-verbally to information for which we lack awareness; with only non-verbal responses, it is difficult to distinguish between conscious and non-conscious processing in other animals. Second, even if we assume that some non-verbal tests reveal aspects of consciousness in non-human animals, the nature of consciousness is likely to be quite different given the human brain's unique capacities for language, hierarchical cognition, conceptualization, prospective cognition and self-reflection, which I believe all contribute to fear and other emotional experiences.
KR: While it is clear that few, if any, animal models fully represent the complexity of human neuropsychiatric disorders, there is tremendous evidence for conservation across species—from mouse to human—for basic behaviors, including for many of the defensive threat responses and their underlying circuits. Data robustly suggest that appetitive and aversive behaviors, respectively, are underlying phenomena for the syndromes of addiction and fear-related disorders such as phobia, anxiety, and post-traumatic stress disorder (PTSD). Furthermore, the subcortical amygdala, bed nucleus of the stria terminalis (BNST), striatal, hippocampal and brainstem circuits, and to some extent aspects of cortical regulatory areas, are highly conserved in form and function across mammals. Decades of work has established a clear circuitry that has largely held up in human imaging and physiology studies and in rodent studies using modern tools such as optogenetics, chemogenetics, calcium and electrophysiology tools. While much more needs to be established, powerful approaches such as single-cell RNA-sequencing across regions and species, large-scale genetic tools combined with transcriptomics, and digital phenotyping across species are enabling truly novel and powerful translational approaches that do not model disorders per se, but instead model their component parts, from molecules to circuits to aspects of behavioral syntax that underlie the defensive ‘threat’ to ‘fear’ continuum.
KT: New technologies and methods can enhance our understanding of fear as they can advance our understanding of brain circuitry and function in general. Fear conditioning is often a first proof-of-principle behavioral paradigm used to validate new technologies because it is so robust and reproducible.
Q7: How can new technologies and methods enhance our understanding of fear?
RA: Much attention has been paid to increasing the precision of measurements and manipulations of the brain, but I think we need to improve the validity of stimuli and measurements of behavior. Only a few studies have used high-dimensional, multivariate measures of behavior. For instance, one can measure the change in the body surface over tens of thousands of little chromatophores that cuttlefish use to camouflage themselves, a measure that has been claimed to give us a direct readout of the animal’s perceptual state. Rich measures in humans would also seem achievable: we need to measure in detail people’s movements in 3D space, their whole-body blood flow and so forth. At the stimulus end, the best stimulus is the real world, and studies in an animal’s natural environment or in a person’s everyday life would help to provide validity to studies in the lab. Virtual reality could probably help here. Of course, behavior isn’t everything (fear doesn’t just function to cause behavior); interactions with other cognitive processes are important to quantify as well. In the ideal case we would probe not only how behavior changes over time when an ecologically valid threat stimulus is presented, but also how this affects memory, attention, perception and decision-making.
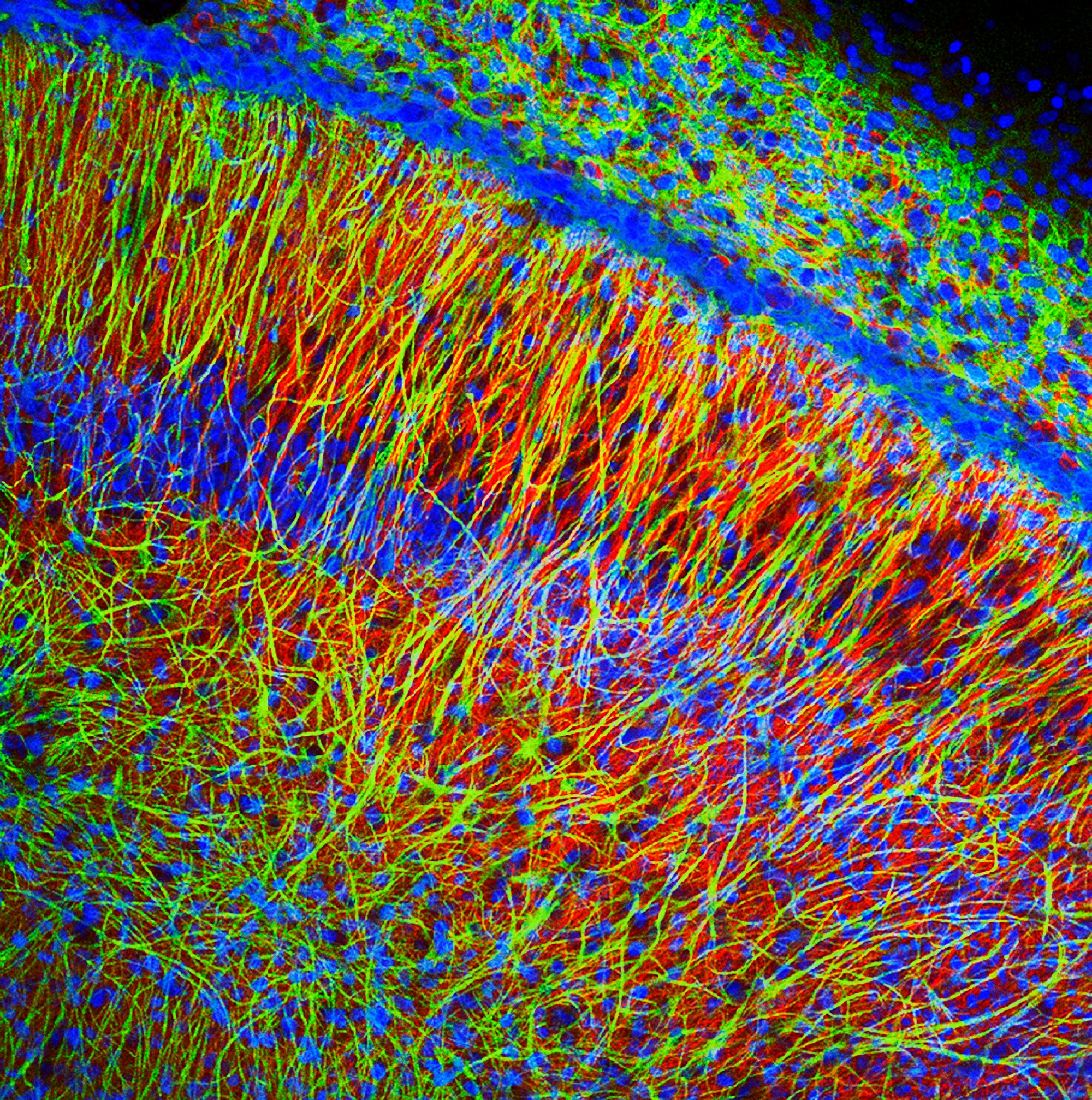
MF: Particularly useful is our ability to map large cellular networks that participate in different situations and behaviors. These have largely been achieved using immediate early gene imaging techniques such as catFISH. Above, I described Bernstein’s research that used this methodology to show that taste aversion and fear conditioning activate largely independent amygdala networks, helping us distinguish two aversive motivational systems. New implantable microscopes also hold considerable promise in advancing our understanding. But our conceptual understanding of phenomena cannot be sacrificed to these technical achievements; the two must advance hand-in-hand.
LFB: New technologies and methods can enhance our understanding of fear by providing the capacity to observe animals in a wider variety of highly variable ethological contexts using higher-dimensional measurement procedures with improved temporal and spatial specificity. The ability to measure and model naturalistic contextual variation is crucial, particularly for genetic studies; most genetic variation related to individual differences that predispose an animal to disease sits in non-coding regions of the genome, which are strongly influenced by context.
JL: The new methods available today are revolutionizing brain research. But sometimes the methods seem to take precedence over the questions. New methods can only help us if we have adequately conceptualized the problems. Complications from poorly designed studies are relatively easily corrected—just do a better experiment. Conceptual problems are harder to change. Ideas become dogma, and dogma typically goes unquestioned; new methods can’t fix that. It’s good that in this exercise we are taking a step back to assess where we are, conceptually, relative to where we need to be.
KR: An array of fantastic new molecular tools, from optogenetics to chemogenetics to in vivo dynamic imaging, has allowed a functional dissection of cells, molecules and pathways that underscore threat processing and inhibition. Understanding these processes will provide novel and robust insights into control of specific kinds of emotional responses, in particular fear and threat. From a translational perspective, such a cellular level of precision of behavioral control leads to remarkable possibilities. Through single-cell RNA-sequencing, we can now assess whether cell types and microcircuits are conserved from mouse to human. Furthermore, we can ask whether these conserved pathways also share molecular targets, so that one could apply data analytics and bioinformatics toward understanding combinations of drugs that might specifically inhibit conserved fear circuits or enhance extinction circuits. For example, even in humans, could we use brain stimulation techniques or even gene therapy to target fear circuits in reliable, therapeutic ways?
Concluding remarks
After this discussion, can we agree on a definition of fear?
RA: I think we want to be careful to leave room open for revision and discovery, rather than rigidly ‘defining’ fear. Perhaps we could agree on these points: (i) fear involves particular regions of the brain, especially clearly subcortical ones. We can measure it from, and induce it by manipulating, particular neural circuits (for example, the amygdala) and not others (for example, the cerebellum). Whether these circuits are specific to fear is a further empirical matter. We could come up with some initial inventory of how strong the evidence is for the participation of particular brain structures in fear. (ii) There are subtypes, varieties or dimensions of fear. I would advocate, in the first instance, for differentiating it based on functional criteria. We could come up with lists here, too. (iii) The state of fear, the conscious experience of fear, the concept of what ‘fear’ means and the meaning of the word ‘fear’ are all different things (the latter two can only be studied in humans). If you give people words or stories to rate, you are testing the last two. It would be useful to come up with taxonomy or a glossary for this.
MF: Several of the approaches (Aldolphs, Ressler, Tye and Fanselow) seem to take evolutionary concerns and commonalities between fear expression as central. Importantly, these approaches recognize that something can be learned from all measures of fear. LeDoux and Feldman Barrett stand apart. In my opinion, their approaches suffer from the human tendency to glorify verbal report over all other measures. So, the hurdle is to agree to treat verbal report as informative, but not exclusively so. LeDoux’s description of the circuitry supporting conscious reporting of fear recognizes that there is significant input from the amygdala and other components of the antipredator system. I believe this is also true of Feldman Barrett’s description, although she does not discuss explicit circuitry. The circuitry that gives rise to any individual fear response will have two components. One component arises from the core defensive circuit, and this will be similar for all fear responses. But there will also be a second component providing specific information, and the processing necessary, for execution of the particular response. This is just as true of freezing as verbal report. Each response will have its own unique subcircuit, part of which will belong to an essential circuitry common to all fear responses. Each response reflects both fear and other contextual information. If we recognize this, then we may be close to consensus. Even something seemingly simple as freezing is a complex construction. The firing of basolateral amygdala neurons that initiates freezing is brief and transient and needs to be converted elsewhere into the firing patterns necessary to maintain a sustained motor response. The motor pattern we call freezing varies considerably in posture; the freezing rat can be crouching on the ground or rearing up and leaning on a wall. This is remarkably similar to Feldman Barrett’s description of ‘many to one’ response mapping where the ‘intention’ to freeze is implemented by different motor plans. Freezing does not occur in random places: animals preferentially freeze near walls, in corners and in dark locations. Thus, the freezing subcircuit processes visual contextual information that is quite separate from the sensory stimuli that signal danger. Past experiences will also influence current action. These multiple streams of information must coalesce in a manner that supports each instance of freezing. Thus, even freezing is, in Feldman Barrett’s words, “highly context-dependent and variable.” Maybe we are not so far apart after all.
LFB: I am optimistic and hopeful that scientists can reach agreement on defining fear, but it will require that we reconsider some of our ontological commitments and the philosophical assumptions that ground our empirical inquiry. Several of the debates within the science of fear (and the science of emotion, more generally) are philosophical rather than scientific and so are unlikely to be resolved with experiments or data. Still, discussions like these are worth having, because commitments and assumptions are conceptual tools that influence (and constrain) the process and products of scientific inquiry.
JL: The fundamental issue we are discussing is the role of subjective experience in the science of emotion. Is it one of many aspects of emotion, or is it what emotion is all about? This is a perennial issue in emotion theory. The reason we are discussing this as if it was a novel topic here is because much contemporary research on the brain mechanisms of fear has involved fear conditioning, which has largely been isolated from mainstream emotion theory. My PhD dissertation in the late 1970s included studies of emotional consciousness in split-brain patients and introduced me to the cognitive theory of emotion. Ever since, I have viewed emotions as cognitively assembled states and tried to integrate cognitive thinking about emotion into the ‘fear’ conditioning (or what I call ‘threat conditioning’) field. But it has been an uphill battle. For example, sometime in the late 1980s, one of my colleagues from the behaviorist tradition asked me, “why do you talk about fear conditioning in terms of emotion?” These days, for better or worse, emotion talk is fairly common in the animal aversive conditioning field. But the conception of emotion is often still heavily influenced by the Miller–Mowrer behaviorist ‘fear theory’ from the 1940s, which treated conditioned ‘fear’ as the underlying factor in avoidance. While some from the behaviorist tradition, especially in the tradition of Tolman, viewed fear in animals as an intervening varaiable, a hypothetical ‘central state’ (for example, a hypothetical nonsubjective psychologicial or physiological state) that might connect stimuli with behavior, others viewed it as a subjective conscious experience; however, most did not take a stand either way, which has engendered much confusion. Research on the brain mechanisms of fear in humans has also often used the term ‘fear’ in ways that conflate behavioral and physiological responses with subjective experiences, further adding to the confusing state of affairs in which now find ourselves. As I noted above, some of the disagreements among the participants in this discussion are mostly semantic. But, also as noted, semantics are crucial to our conceptions and assumptions. It’s a good thing that different ideas are being expressed. Fear has too long been talked about in ways that imply we all mean the same thing. Now that different conceptions are being openly discussed, it would, as I suggested above, be useful for researchers to be more rigorous and vigilant in defining what each means by ‘fear’ each and every time the term is used, so that others will understand what is being referred to in a given instance. The less cumbersome alternative, which I prefer, is simply to confine fear to fear itself. As the social psychologist Matthew Lieberman recently argued, “emotion is emotional experience”. More generally, mental state terms like fear should be used to refer to mental states and not to behavioral or physiological control circuits.
KR: I believe that we can agree on a definition. I think most everyone already states some of the shared understanding of a subset of the conscious awareness components in humans, as well as observable physiological and behavioral components in humans and model systems. I think that separating the salience, valence and action (or perhaps feeling, perception and behavior) descriptions will help with some of the semantics. Additionally, I think that focusing on pragmatism over theoretical will help with efficiency toward a workable definition.
In your view, what are the clinical implications of a clear definition of fear?
RA: The clinical implications are huge. Probably the best evidence for this is the paper by LeDoux and Pine, and subsequent rebuttals by Fanselow. LeDoux and Pine argue that the effects of anxiolytic drugs studied in rodents do not inform about the conscious experience of fear and that this is why anxiolytic drugs don’t work well for alleviating fear in humans: they are aiming at the wrong target. For instance, an antidepressant that makes depressed people really awake and active and gets them out of bed in the morning would not be helpful if they still feel depressed. This is just one example, but it shows how important it is to figure out what we are studying when we study fear in animals and in humans and when we measure or manipulate its neural components.
MF: The scientific definition of fear must help us understand the clinical manifestations of fear. Let’s start with what I see as the two big questions. First, why are anxiety disorders so prevalent? Elsewhere I’ve described this as a natural and predicted consequence of the costs and benefits of hits vs. misses when assessing the presence of threat. Second, why are anxiety disorders so detrimental? Fear, anxiety and panic in the absence of actual danger are not beneficial, so why doesn’t the realization of this fact make anxiety disorders disappear? I believe this is a consequence of engaging a system whose strategies are determined by contingencies that operated over phylogeny rather than ontogeny. I also come back to my point that if consciousness evolved to allow flexible and rational decision making, the lack of flexibility and rational action that characterizes anxiety disorders suggests that conscious contributions are limited. I’m not saying that there is no contribution, but we must temper our conclusions with the facts of the clinical situation.
LFB: One goal of understanding the neurobiological basis of fear is to aid the treatment and prevention of mood-related symptoms in both mental and physical disorders. This goal will be accomplished only when we consider the mechanisms and features of fear in the context of what the broader range of evidence actually suggests about the evolution and development of the nervous system. An evo-devo approach requires considering what the broader range of evidence actually suggests about features of the human nervous system that are deeply evolutionarily conserved vs. features that emerge during human vs non-human brain development. In addition, scientists should understand that disorders which strongly implicate fear and/or anxiety, such as PTSD, are not specific ‘fear’ disorders; this has implications for how these disorders are understood, treated and prevented.
JL: In the face of a sudden danger, we typically consciously experience fear and also respond behaviorally and physiologically. Because the experience and the responses often occur simultaneously, we have the sense that they are entwined in the brain and thus are all consequences of a fear module. This is a common and popular view of fear, and it has led to search for medications and behavioral treatments that will relieve subjective distress in patients suffering from fear or anxiety disorders. Since the behavioral and subjective responses are both assumed to be products of a fear module, it is also assumed that treatments that alter behavior in animals will alter fear and anxiety in people. Few would claim that this effort has been a rousing success. Small but statistically significant differences relative to placebo controls are found in some studies, but for any one individual the chances of successful treatment are much lower than desirable. And even when successful, side effects pose other problems. But more pertinent to our concern here is why these treatments help, when they do. Is it because the treatment directly changes the content of the subjective experience, or because it indirectly affects the experience (for example, by reducing brain arousal, feedback from body responses), or because it affects cognitive processes that contribute to the experience (episodic and semantic memory; hierarchical deliberation, working memory, self-awareness), or all of the above? For the patient it probably doesn’t matter how a treatment works, but for the purpose of finding new and better medications, knowing the underlying mechanism of action is crucial. And to understand this we need a conceptualization of not just how the brain controls behavioral and physiological responses elicited by threats, but also how the threat engenders the conscious experience of fear—something that can only be explored in humans. After many decades of being marginalized as ‘just another measure of fear’, there is renewed interest in consciousness (including emotional consciousness) in psychology, neuroscience and the various psychotherapeutic communities—not simply because subjective experience is an interesting research topic, but also because it plays a central role in our lives and must be a central part of therapy.
KR: Disorders of fear processing (and related panic and anxiety), from panic disorder, social anxiety and phobia to PTSD, are among the most common of psychiatric maladies, affecting hundreds of millions of people worldwide. Combined, they are also among the highest in terms of morbidity, loss of work, comorbid psychiatric and medical disorders, and mortality from suicide. Despite these unfortunate statistics, we understand these disorders moderately well and have reasonable treatments. These disorders all share the core emotion of fear and threat-related symptoms. The diagnosis of a panic attack, shared among all of these disorders, includes racing heartbeat, sweats, chest pains, breathing difficulties, feelings of loss of control and a sense of terror, fear, impending doom and death—basically the ‘fear reflex’ run amok! The reflexes and symptoms that are ‘normal’ in a threatening situation are experienced by those with anxiety disorders all the time—as if they can’t ‘turn off’ the fear switch. Furthermore, the most well-supported, empirically validated treatments for these disorders rely on repeated exposure, now understood as the process of ‘fear extinction’. Advances in our understanding of mechanisms of fear and threat-processing, its underlying neural circuitry and molecular biology, and improved methods of fear inhibition and extinction, will contribute to advancing treatment and prevention for these devastating disorders.
What is an important gap that future research (and funding) should try to fill?
RA: Integrative, cross-species research. Right now, research on fear (and other emotions) is like the blind men and the elephant. Each lab studies either humans or a single animal model, and each study focuses on a narrow aspect of fear. We need to figure out how to put all this together. I’m not suggesting a giant project where all manner of species and humans are studied, but we should produce standardized sets of experimental protocols that the scientific community can use—in particular, these protocols and their measures have to cut across species to some extent. Right now, research on fear in animals and in humans is really disconnected, and that has to change if we are to make progress. We need uniform criteria for evaluating papers and grants and for building a cumulative science of fear. Needless to say, the by-now-common criteria of reproducibility and data sharing should apply also.
MF: Current technical developments in neuroscience are both important and breathtaking, but where we fall short is conceptual development and advancing formal theories of behavior. Without conceptual development, the data being collected with those tools can be, and often is, profoundly misinterpreted. While some of the contributors to this discussion bemoan the influence of behaviorism, I feel that a far more problematic trend is the intuitive, and often anthropomorphic, approach to behavior that characterizes much of the most technically advanced neuroscience going on now. This caution was a major motivator for the initial development of behaviorism. Again, I note that the negative comments regarding behaviorism above were directed at an outdated form of behaviorism that learning theorists discarded decades ago, and these comments can therefore be considered strawman arguments. Behavior is of paramount importance, not only because it allows objective observation, but also because it is where the organism connects with selection pressure. Careful observation of emotionally charged animals shows that behavior is often irrational and our intuitions about how to interpret it are likely to fail. I call ‘predatory imminence theory’ a functional behavioristic approach because its ideas flow from concerns about both evolution and behavioral topography.
LFB: Every behavior is the result of an economic decision about an animal’s global energy budget and involves estimating expenditures and deposits over various temporal windows that are relevant to the niche of the animal, taking into account the animal’s current physiological condition. If fear is to be understood in an evolutionary and developmental context, then it must be studied in the reality of those economic decisions as they emerge in an animal’s ethological context. More attention must be paid to basic metabolism and energy regulation, including the cellular respiration of neurons and glial cells. A predictive processing approach, rather than a stimulus–response approach, must also be considered. And a greater emphasis on variation and degeneracy, at all levels of analysis, as well as neural reuse, must be considered.
JL: My view is that the biggest impediments to progress are our conceptions and the language we use to characterize psychological constructs. My personal preference is that mental-state terms, such as fear, should be avoided when discussing relatively primitive processes that control behavior; mental state words should only be used when specifically referring to mental states, such as the conscious experience of fear.
KR: I agree with Tye that “given its critical importance in survival and its authoritarian command over the rest of the brain, fear should be one of the most extensively studied topics in neuroscience, though it trails behind investigation of sensory and motor processes due to its subjective nature.” I feel that it is among the ‘lowest hanging’ fruit in behavioral and translational neuroscience, and that an explanatory science—from molecules to cells to circuits to behavior—will provide a transformative example for other areas of neuroscience and neuropsychiatry. I think current gaps include many of the questions raised in this discussion, such as how are valence, salience, perception and action separated at a neural circuit level. Are there critical differences between predatory vs. social survival circuits and between reactive vs. cognitive fears? How discrete, at a cellular circuit and microcircuit level, are the different components and behaviors underlying threat processing? Finally, from a translational perspective, how are the molecules, cells and circuits conserved in humans—which ones constitute convergent evolution of similar behaviors with distinct mechanisms vs. which represent truly conserved mechanisms that are essentially the same in rodents and humans?
KT: The field would benefit greatly from additional paradigms that are distinct yet stereotyped to facilitate the same critical mass of research surrounding it that Pavlovian fear conditioning has undergone to really be able to make comparisons.
Summary
Substantial progress has been made in our understanding of the neural circuits involved in fear. This has been a cross-species endeavor, yet—as debated here—there are disparities on how to investigate and define fear. We hope that the debate presented here, which represents the views of a subset of outstanding researchers in the field, will invigorate the community to unify on clear definitions of fear (and its subtypes) and to show the courage to pursue new behavioral assays that can better differentiate between fear circuits (or concepts) involved in perception, feeling and action. The implications will be far-reaching, as a lack of coherence on what neural systems are involved in fear and fear learning will hinder scientific progress, including the study of human affective disorders such as PTSD, anxiety and panic disorder. That is, how we define fear determines how we investigate this emotion.
This article is reproduced with permission and was first published on July 22, 2019.
